§ Islet of Langerhans in pancreas
一.在這一節中,我們會透過將Islet of Langerhans中的細胞分成三種來介紹每一種細胞會分泌的hormone。先來用一張Netter手繪的圖來幫大家複習一下組織學:

Cell type
|
Hormone
|
α cell(25%)
|
Glucagon(升糖素)
|
β cell(65%)
|
Insulin(胰島素)
|
δ
cell(10%)
|
Somatostatin(體抑素)
|
(一)α cell & glucagon
1.Glucagon的結構:

2. Gucagon 的主要target cells:

(1)Glucagon主要的target cell有adipose tissue上的adipocyte,以及liver裡的
hepatocyte。
3.Glucagon在target
cell中的生化反應機制:

cytoplasm側有黏上一個由α,β,γ三個subunit所組成的G protein。值得一提的是,在
這個G protein的α subunit上,又另黏有一個GDP,會在接下來的反應中扮演重要角
色。
(2)當glucagon接上glucagon receptor, 這個由glucagon receptor和G protein構成的
complex會發生結構上的改變,將原本包在結構裡的GDP露出來,讓cytoplasm裡
的GTP能夠交給它一個磷酸根,使黏在α subunit上的GDP變成GTP。
(3)當α subunit上的GDP變成GTP時,α subunit會脫離整個complex,去用自己身上
GTP的磷酸根活化adenylate cyclase.
(4)當adenylate cyclase接上一個來自α subunit上GTP的磷酸根而活化時,它會轉化
cytoplasm中的ATP成為cAMP。而這個cAMP,會進一步的引發PKA pathway(
Protein Kinase A pathway)。
4.Glucagon 在不同target cell引起的後續反應:
之:
Type of target cell
|
Reaction
|
Result in plasma
|
Hepatocyte
|
促進:glycogenolysis, lipolysis
抑制:glycogen synthesis,
glycolysis
|
Plasma
glucose增加
Plasma
FFA增加
Plasma
glycerol 增加
|
Adipocyte
|
促進:lipolysis
抑制:fatty acid
synthesis
|
Plasma
FFA增加
Plasma
glycerol 增加
|
促進
|
抑制
|
|||
Neural
transmitter
|
Hormone
|
Neural transmitter
|
Hormone
|
產物
|
β-adrenergic stimulator
acetylcholine
|
cortisol
CCK
|
GABA
|
Insulin
Somatostatin
|
Glucose
Ketones
Free
fatty acid
|
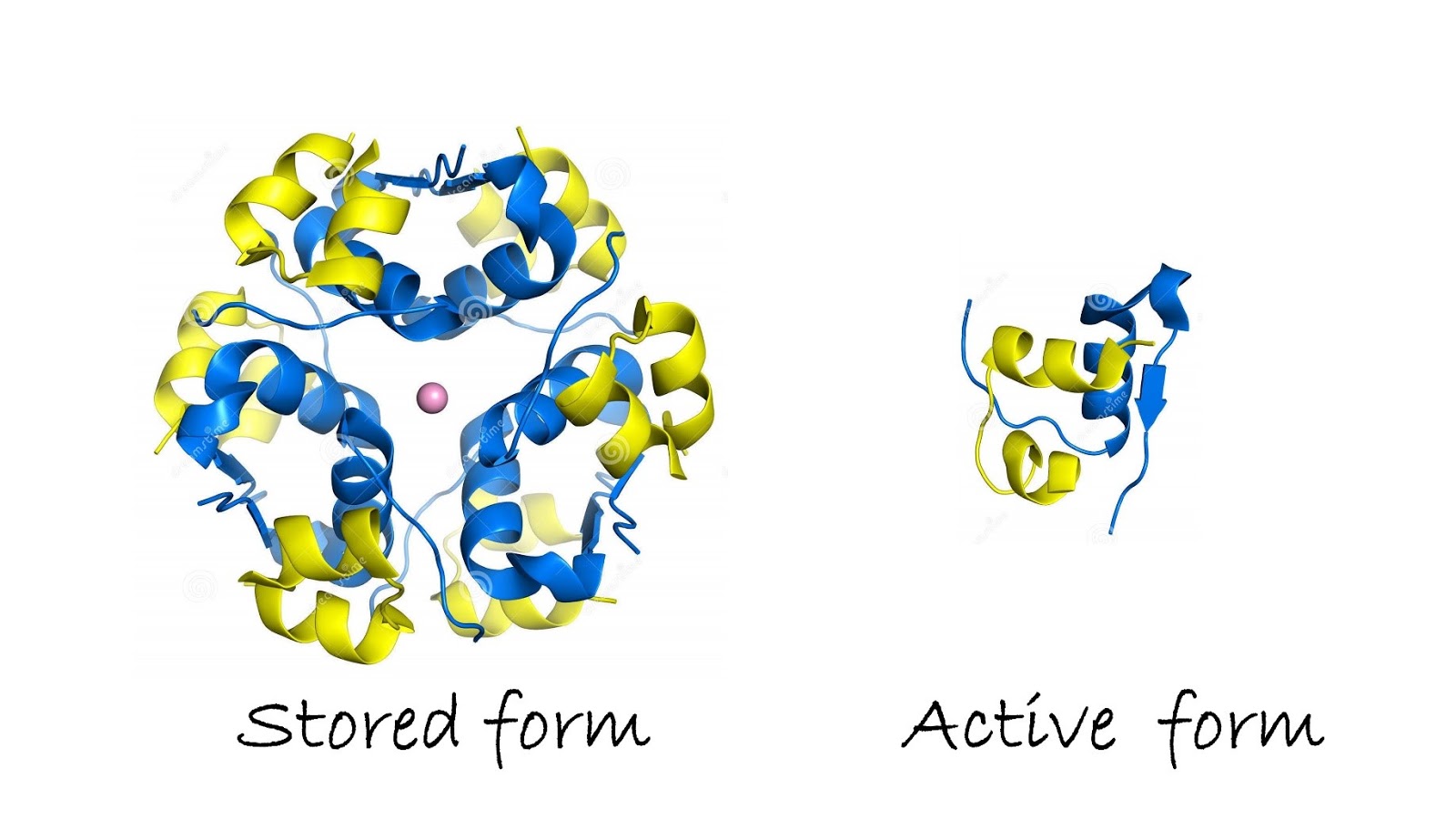
會以一個由Zn2+為中心聚合而成的hexamer儲存在β cell中,等到β cell受到刺激,
才會以monomer的active form釋放出來。
2.Insulin的生化合成機制:

disulfide bond formation enzyme,使A,B chains間形成雙硫鍵,將整條多肽鏈變成
折疊態的proinsulin。接下來,rough ER會將摺疊好的proinsulin送進golgi 裡面,並
在golgi裡面切掉原本用來連接A,B chain的C peptide,形成mature insulin,並連同
C peptide一起儲存在vesicle中,以待刺激來時釋放到β cell外。
已確診的糖尿病患者常有在進行insulin注射的治療,若要直接以其血液中insulin
濃度,來判斷患者β cell的功能,就會被注射進去的人工insulin所干擾。而C-
peptide這個1:1的特性,恰好使它成為臨床上很好的測量替代物。

liver裡面的hepatocyte.
4.insulin在target cell中引起的生化反應機制:

(2)當insulin receptor接上insulin,它就會發生結構上的改變,將自己tyrosine kinase
domain上的tyrosine做自我磷酸化。這個經過自我磷酸化的insulin receptor會吸引
位於cytoplasm中的ATP和IRS(insulin receptor substrate)黏到β subunit上面。
(3)當兩個ATP和IRS都黏到β subunit上面之後,β subunit就會從兩個ATP身上各拿一
個磷酸根黏到IRS上去活化它。
(4)這個被活化的IRS會直接去啟動PI3K pathway,或者是藉由活化RAS蛋白去啟動
MAPK pathway.
5.Insulin在不同target cell引起的後續反應:
PI3K pathway和MAPK pathway會在不同細胞中引起不同的反應,表列如下:


(1)先用一張表格整理之:
促進
|
抑制
|
|||
Neural
transmitter
|
Hormone
|
Neural transmitter
|
Hormone
|
Product
|
Acetylcholine
|
Incretin
|
α2-adrenergic agonist
β-adrenergic agonist
|
Somatostatin
Leptin
|
Glucose
|
接下來會分別說明這些物質對insulin調控機制。
(2)含有insulin 的vesicle分泌機制:vesicle出去,所以我們必須來檢視一下它們的分泌機制:

channel;含有insulin的vesicle也必須以synaptobrevin 2黏在syntaxin 1上(這兩個
也可以換成任何功能類似的東西)。
(b)當voltage-gated Ca2+ channel附近的cell membrane出現去極化後,會讓voltage-
gated Ca2+ channel打開,Ca2+也會順著濃度梯度流進細胞裡面。
and chain)就會堵住通道,使整個channel變成不活化的狀態(inactivation)。同
時,流入細胞裡的Ca2+,也會去接上synaptobrevin 2。
(d)接上Ca2+的synaptobrevin 2會發生形變,將含有insulin的vesicle推出細胞外。
(e)被排到細胞外的insulin hexamer 會被水分子以hydrogen bond拉得四分五裂,最
後變成具有活性的monomer狀態。
(3)Acetylcholine--Stimulating neural transmitter
先來看一下acetylcholine的結構:

接下來就來細看acetylcholine如何促進insulin的分泌:

(a)和glucagon的receptor一樣,acetylcholine在分類上屬於G protein coupled
receptor。當receptor接上acetylcholine時,α subunit上的GDP會被變成GTP,進
而暫時脫離整個receptor,跑到旁邊去磷酸化phospholipase C。
(b)這個phospholipase C被磷酸化後,會用它剛得到的磷酸根再去磷酸化一旁的
PIP2(Phosphatidylinositol 4,5-bisphosphate),使它成為PIP3。
(c)當PIP2成為PIP3後,會變得不穩定,自己從細胞膜上脫落,變成PI3。
(d)這個PI3會跑去黏上位於endoplasmic
reticulum膜上的PI3 activated
Ca2+
channel。這個通道就會打開,釋放出ER中儲存的Ca2+到cytoplasm裡面。而前
面我們已經說明過為何[Ca2+]升高可以促進insulin的分泌,此處便不再多提。
先來看一下incretin(GLP-1)的結構:


(a)最一開始的步驟跟glucagon在target cell裡的反應幾乎一模一樣,都會從G
protein活化adenylate cyclase 產生cAMP
(b)接下來,cAMP會接上PKA的regulatory subunit,並讓整個蛋白質結構發生形
變。
(c)發生形變後,PKA的catalytic
subunit就會和regulatory subunit分開。
(d)這個PKA的catalytic
subunit接到位於含有proinsulin之vesicle膜上的
VGLT3(vesicular glutamate transporter 3)。同時,位於vesicle膜上的H+pump也
會持續的將H+從cytoplasm中打進來。(註:CPE=carboxypeptidase
E)

(e)當VGLT3接上PKA的catalytic subunit之後,會將glutamate打進細胞裡。
(f)奇妙的是,這些glutamate被打到細胞裡後又會急著想從GLT2(glucose
cotransporter)跑出去。由於它是cotransporter,出去的時候會夾帶3個Na+和1
個H+到cytoplasm裡頭。

(g)當GLT2打開的時候,K+會搭GLT2的便車從cytoplasm裡頭跑到vessicle裡面來。
(h)由於四個正電荷進去,一個正電荷出來,造成vessicle裡的電位較原本的低,這
也就促進了H+ pump的加速運作,將更多的H+打進細胞裡。

(i)這些不斷累積的H+使得vessicle的pH值降低,進而使儲存在vesicle中的
CPE(carboxypeptidase
E)去和proinsulin結合,並將C peptide給切下來,將A-B
chain釋放到vesicle中,成為mature insulin.
chain釋放到vesicle中,成為mature insulin.
(j)當insulin越累積越多,就會被原本就儲存在vesicle裡的Zn2+所吸引,最後聚合
成 stored form的 insulin hexamer,等待接受刺激釋放出β cell外。
(5)α2-adrenergic agonist(stimulating)
&
β-adrenergic agonist(inhibiting) neural transmitt
先來看一下α2-adrenergic agonist和β-adrenergic agonist的結構:

可以看到,它們其實就是一群有帶著類似epinephrine和norepinephrine的物質。
接下來就來看看α2- & β- adrenergic agonist如何影響insulin的分泌

pathway的底下特別註記了一個”αβ”,原因是雖然他們和incretin一樣是用PKA來
反應,它的總效應卻是抑制!
至於這一切怎麼發生的,讓我們繼續看下去…

(b)(5)當catalytic PKA被從cAMP上切下來之後,它會跑到細胞核裡面去和跨騎在
insulin gene上的CREB(cAMP response element-binding protein)結合,並活化
它,讓它開始進行insulin的transcription。
(c) (6)同時,也有部分的PKA會去黏上細胞膜表面的K+ ATP channel,並促進這個
channel Kir6.2 subunit的磷酸化。一旦這個Kir6.2 subunit被磷酸化,就會打開通
道,利用主動運輸的方式把K+從cytoplasm送到細胞膜外。這樣的過程會造成
細胞內帶正電離子的流失,產生過極化,而前面說過,含有insulin的vesicle要
靠細胞膜的去極化才能分泌出去,故PKA的這個作用會抑制insulin的分泌。
(d) 我們稍微思考一下便可以知道,雖然PKA能促進insulin的新合成,但是分泌不
出去,生產再多都沒用,故整體來說是抑制的效果。
(6)Somatostatin—inhibiting hormone
先來看一下它的結構:

可以發現它是一個peptide hormone。
接下來我們直接來看somatostatin如何影響insuliin的分泌。

(a)(1~4)可以發現,它其實和glucagon在target cell中的反應一樣,都是透過G-
protein去啟動PKA pathway。

(b)當PKA的catalytic
subunit被切下來時,它會去黏在GIRK channel(G-Protein-Gated
Inwardly
Rectifying Potassium Channel)和K+
ATP channel的Kir6.2上面,使這兩個
通道打開,讓K+從胞外流向cytoplasm裡面,造成胞內膜電位的下降,引起過極
化,抑制insulin的分泌。
(7)Leptin—inhibiting hormone
先來看一下它的結構:

接下來我們直接來看leptin 如何抑制Insulin的分泌。

(a)和insulin在target
cell中的反應一樣,leptin所用的是會在兩側各接上一個
JAK3(just another kinase 3),具有tyrosine kinase domain的leptin receptor(Ob)。
(b)當leptin接上leptin receptor後,receptor的tyrosine domain會被自我磷酸化,讓游
離在cytoplasm中的兩個STAT3(Signal Transducer and
Activator of transcription 3)
和兩個ATP黏到receptor上去。
(c)當兩個ATP和兩個STAT3都連接receptor上後,β subunit和JAK3就會把兩個ATP 上的磷酸根拔下來各裝一個到兩個STAT3身上。
(d)當兩個STAT3都各接上一個磷酸根後,會從receptor上脫落到cytoplasm中。

(e)這兩個磷酸化的STAT會結合成dimer,並從cytoplasm裡面跑到nucleus裡面,黏
上並阻斷insulin gene的transcription。
(8)Glucose—inhibiting product
Glucose的構造太簡單,我就不放圖了,直接來看它對insulin分泌的影響。

(a)當plasma glucose的濃度變高時,它會從GLUT2(glucose transporter 2)這個通道
跑進β cell裡面。
(b)由於跑進β cell的glucose增加許多,它們就會以勒沙特略原理加強糖解作用
(glycolysis)的進行,因而產生許多ATP。
(c)這些增加的ATP會去活化細胞膜上K+ ATP
channel,使它將更多位於cytoplasm
中的K+打出細胞外。而K+的流失會造成過極化,抑制insulin的分泌。
7.Insulin 和血糖變化的一些直接證據
講完了insulin的生化反應,我們再來看一些較為表淺的現象增加insulin對血糖變化
影響的印象。
(1)Biphasic
response and insulin secretion

(a)這裡要先提出一個問題:insulin是平常就會產生,還是等到接受刺激後才產生
呢?
(b)為了解答這個問題,科學家將glucose快速且持續的注射進受試者的體內。
(c)如果insulin是接受到刺激後才產生的,那它的濃度應該會像上圖左邊的假想曲
線一樣,從濃度0緩慢上升。然而根據實際測到的數據來看,insulin 的濃度會
先衝上來,然後又快速掉下來後,再緩慢上升。
(d)其中的一種解釋方法,是β cell會先儲存一部份insulin在cytoplasm的vesicle裡
面。當glucose急速上升時會先拿這些存在vesicle裡的insulin來應急,等到這些
insulin用完了,再啟動insulin的新生反應。
(2)Incretin
and insulin secretion

(a)這個實驗其實就是前面incretin對insulin分泌有促進反應的一個證明。科學家將
受試者分成兩組,一組口服glucose,一組靜脈注射glucose,在之後的180分鐘
裡維持兩組受試者plasma glucose的濃度,並觀察他們各自血中insulin的濃度。
從右邊experiment的圖表中可以看見,口服glucose的受試者plasma glucose會比
較高,說明當glucose經過腸道細胞時,會刺激腸道細胞分泌incretin,增加
insulin分泌的量。
(b)不過這裡延伸出一個小問題:為什麼科學家要維持glucose的恆定呢?
要回答這個問題,我們必須做兩個假想實驗。
(1)每段時間裡攝入固定量的glucose
我們必須先做兩個假設:insulin在和receptor反應後不會消失,而是會繼續在血
液中存在;每攝入dM的glucose,就有一定量的glucose會因為insulin的效應而
被抽離到血液之外,但是這個抽離的量不會大於dM。

實驗一開始的insulin濃度為0。當在dt的時間裡給予受試者dM的glucose時(1),
會使plasma裡的insulin上升(2),其中,由於口服glucose會有incretin的促進效
應,故其insulin濃度上升得比較多。而由於口服glucose組plasma
insulin上升的
比例比靜脈注射組來得多,在接下來的血糖變化上(3),靜脈注射組的血糖濃
度上升幅度就會比口服組的來得高。接下來的步驟就會出現問題了(4),因為
我們現在知道由於口服glucose組的血糖濃度比靜脈注射組的來得低,它insulin
上升的幅度應該也要比較靜脈注射組的來得低。然而,如果考慮到incretin對
insulin的促進效應,我們就很難預測口服glucose組血糖上升的幅度是多少、會
不會超過靜脈注射組的,因此,固定plasma glucose的攝入量,並不是一個好
的實驗,因為這樣會需要同時考慮兩個因素,造成實驗解釋上的困難。
因此,我們來進行第二個假想實驗:
(2)固定血糖濃度:

由於現在血糖濃度是完全由人工控制的,因果關係變成非常單純的單項關
係,也就是說,glucose的濃度會去影響insulin的分泌,但是insulin的濃度不會
影響到glucose的濃度!所以從圖中各步驟中可以看到,incretin對 insulin分泌的
影響效應隨著glucose濃度的升高很好的顯現了出來!所以我們得到一個很重要
的結論,那就是這個實驗中的plasma
glucose濃度,不管是上升或下降,都是
由一台機器所控制的!
(三) Somatostatin與δ cell
由於somatostatin的結構在insulin和somatostatin的互動關係中已經提過,且它的作用主
要是去抑制各種hormone,和血糖的上升下降並沒有直接的反應關係,故在下面直接
表列它的作用,還有調控它分泌的一些因子。
1. Somatostatin的功能
它會去抑制:
Endocrine
|
Exocrine
|
Neural transmitter
|
Growth hormone
Thyrotropin
Insulin
Glucagon
|
Gastric acid
pepsin
|
Neuron on intestinal smooth muscle
|
促進
|
抑制
|
|||
Hormone
|
Neural transmitter
|
Product
|
Hormone
|
Neural transmitter
|
各種gastrointestinal hormone
glucagon
|
β-adrenergic
cholinergic
|
Glucose
Amino acids
FFA
|
Insulin
|
α-adrenergic
|
台大醫學05生理共筆
還有沒寫上去的數篇paper(以後再補)
沒有留言:
張貼留言